Hybridization in closely related grasshopper species
Hybrid zones, where genetically distinct
groups of organisms interact, leaving at least some offspring of mixed ancestry, are found
in all major groups of the higher organisms (Barton and Hewitt, 1989; Harrison, 1986,1993;
Jiggins and Mallet, 2000). Hybrid zones are supposed to be natural laboratories that offer
experimental material for studies of the characters and processes involved in divergence
and speciation. Hybridizing taxa may be at a crucial stage of speciation when reproductive
isolation between them is incomplete. The formation of new species must involve the
evolution of barriers to gene flow within existing species, but how these barriers arise
remains a problem. Among pre-mating barriers, assortative mating seems to play a primary
role in speciation. Assortative mating can be a by-product of changes in habitat use or
temporal isolation. More intriguing processes occur when assortative mating evolves as a
result of sexual selection when competition over mates in many different forms (like
contests and mate choice) takes place (West-Eberhard, 1983; Andersson, 1994). Divergence
of mating signal systems can also arise via reinforcement, in which natural selection acts
to reduce the production of unfit hybrids (Butlin, 1987).
More research studies were focused on
post-mating barriers when hybrids appeared to be of less fitness than parental species.
Hybrid fitness can be reduced by 1) sterility and inviability; 2) disruptive ecological
selection; 3) disruptive sexual selection. Most attention has been focused on the first
effect, hybrid sterility and inviability, with the majority of studies using Drosophila.
In many other species that are hybridized in nature, hybrids were shown to be either
inviable in one or in both directions of cross, or F1 males were sterile (Hewitt et al.,
1987; Harrison and Bogdanowicz, 1997; Butlin, 1998). Rather less experimental studies were
done in investigation of disruptive sexual selection. Several recent examples have shown a
reduced mating probability of F1 hybrids and preference for conspecific over F1 hybrid
males (Naisbit et al., 2001). 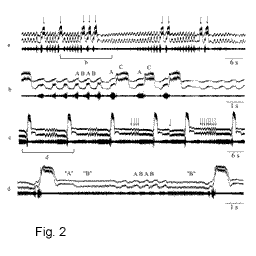
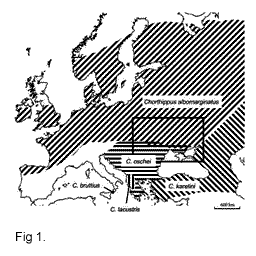
Four sibling grasshopper species - Ch.
albomarginatus, Ch. oschei, Ch. karelini and Ch. lacustris - have
similar calling songs but extremely different and elaborate courtship songs, which were
described by v.Helversen (1986). Besides singing, three species (Ch. oschei,
Ch. karelini and Ch. lacustris) demonstrate spectacular movements of the
legs, palps and antennae during courtship (v.Helversen, 1986). The species are
geographically separated over much of their ranges (Fig. 1).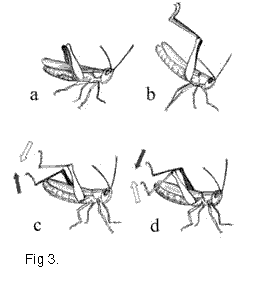 On the basis of analysis of the courtship songs and stridulatory movements of the hind
legs, a wide hybrid zone between two species Ch. albomarginatus and Ch.
oschei has been revealed (Vedenina and v.Helversen, 2003). In 15 localities studied
in Ukraine and Moldova, not only pure species (Fig. 5)
but also individuals with intermediate song characteristics in terms of temporal
parameters of the song and leg-movements have been found. The specimens singing strictly
intermediate pattern were in minority, most of the specimens sang the songs that resembled
a parental type. However, for most of the songs recorded in the hybrid zone, a high degree
of inter- as well as intra-individual variability was found (Fig. 2). Some hybrid males
produced very conspicuous up and down movements with the hind tibiae in the beginning of
the C element (Fig. 3 c,d, 4d). These movements can be regarded as a new element of the
courtship behaviour that is not known for either of the pure species. A comparison of the
leg-movement patterns in hybrid grasshoppers allowed a suggestion to be made about the
homology of sound elements in the albomarginatus and the oschei songs (Fig. 4). For
example, the elements A1 and B1 in the oschei song seem to originate from the A and B
elements correspondingly.
On the basis of analysis of the courtship songs and stridulatory movements of the hind
legs, a wide hybrid zone between two species Ch. albomarginatus and Ch.
oschei has been revealed (Vedenina and v.Helversen, 2003). In 15 localities studied
in Ukraine and Moldova, not only pure species (Fig. 5)
but also individuals with intermediate song characteristics in terms of temporal
parameters of the song and leg-movements have been found. The specimens singing strictly
intermediate pattern were in minority, most of the specimens sang the songs that resembled
a parental type. However, for most of the songs recorded in the hybrid zone, a high degree
of inter- as well as intra-individual variability was found (Fig. 2). Some hybrid males
produced very conspicuous up and down movements with the hind tibiae in the beginning of
the C element (Fig. 3 c,d, 4d). These movements can be regarded as a new element of the
courtship behaviour that is not known for either of the pure species. A comparison of the
leg-movement patterns in hybrid grasshoppers allowed a suggestion to be made about the
homology of sound elements in the albomarginatus and the oschei songs (Fig. 4). For
example, the elements A1 and B1 in the oschei song seem to originate from the A and B
elements correspondingly. 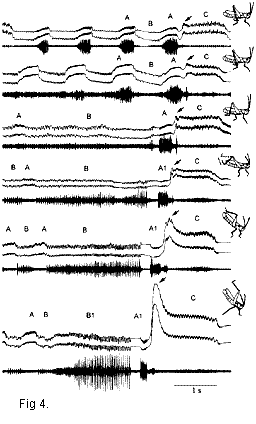
In the laboratory, Ch. albomarginatus and Ch. oschei were hybridized and F1 and F2 hybrids were obtained. The courtship songs of the laboratory hybrids were recorded and compared with the songs of the natural hybrids. The song analysis showed that the natural hybrids were mainly the offspring of the backcrosses (Vedenina et al., 2007). The question arises about the barriers to gene flow developed between two hybridizing species Ch. albomarginatus and Ch. oschei. Behavioural experiments showed that the females of both species were quite selective to conspecific courtship (Fig. 6). Moreover, assortative behaviour is very likely to be based on discrimination of the acoustic signal. One can conceal that these species have developed pre-mating ethological isolation, although an incomplete one, since in 10-20 % of cases the females mated with heterospecific males.
Study of the hybrid fitness showed a
decreased embryonic and larval viability in F1 and F2 hybrid offspring (Vedenina et al., 2007).
Hybrid unfitness was stronger in one direction of cross when Ch. oschei was the
maternal species. Asymmetrical genetic incompatibility is indicative of an incomplete
post-mating barrier evolved between two species. 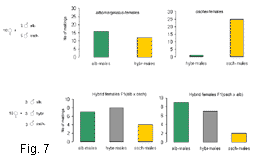
We studied whether the hybrids have a reduced mating probability compared with the parental species. In the behavioural experiments the hybrid females demonstrated much lower selectivity than the females of the pure species. The highest selectivity was found in the females of Ch. oschei: they preferred conspecific males over hybrid males and the males of Ch. albomarginatus (Fig. 7). The females of Ch. albomarginatus did not distinguish between conspecific and hybrid males. The hybrid females did not distinguish between all three types of males (hybrids, Ch. albomarginatus and Ch. oschei). The results evidence that the hybrid females in mixed populations may successively compete with the females of pure species for the mates. In contrast, the hybrid males may successively compete with the males of Ch. albomarginatus but lose to the males of Ch. oschei.
We suppose that the two species are different in habitat preferences in the contact zone. This problem is currently investigated.