Гибридизация у близкородственных видов саранчовых
Проблема гибридизации в
природе между близкородственными видами все
чаще привлекает к себе внимание исследователей.
Гибридные зоны представляют собой уникальный
экспериментальный материал для исследования
процессов видообразования, в частности, порядка
установления изолирующих барьеров (Harrison, 1986; Jiggins,
Mallet, 2000; Butlin, 1998; Mac Callum, 1998). Принято выделять пре- и
посткопуляционные изолирующие барьеры.
Прекопуляционные барьеры могут возникать в
результате установления избирательного
спаривания, которое может быть побочным
продуктом различных экологических (сезонных или
биотопических) изменений. Избирательное
спаривание может также возникнуть как результат
полового отбора (Andersson, 1994). Сравнительно недавно
было доказано, что прекопуляционные барьеры
могут устанавливаться в отсутствии
посткопуляционных, а именнно близкородственные
виды, не скрещивающиеся в природе, могут успешно
гибридизоваться в лаборатории и иметь
плодовитое потомство. Исследованию
посткопуляционных механизмов изоляции - таких,
как гибель гамет и зигот, нежизнеспособность и
стерильность гибридов, посвящено много работ (Orr,
1996; Haldane, 1922; Naisbit et al., 2002). В то же время, гибриды
могут быть фертильны и жизнеспособны в
лаборатории, но проигрывать родительским видам в
приспособленности к конкретным экологическим
условиям или в успехе привлечения полового
партнера (Hatfield, Schluter 1999; Naisbit et al., 2001). Таких
исследований проведено сравнительно немного.
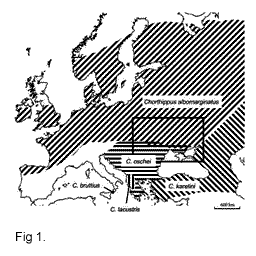
Два европейских
близкородственных вида саранчовых Chorthippus
albomarginatus и Сh. oschei (Orthoptera: Gomphocerinae)
гибридизуются в контактной зоне на Украине и в
Молдавии (Vedenina, Helversen, 2003) (Рис. 1). Виды хорошо
различаются по сигналам ухаживания, которые
чрезвычайно сложны по амплитудно-временной
структуре и, кроме того, у Ch. oschei
сопровождаются характерными демонстрационными
движениями задних ног (Рис. 5).
Анализ сигналов ухаживания особей из гибридных
популяций показал, что во всех исследованных
популяциях особи с промежуточными сигналами
составляют меньшинство, тогда как преобладают
особи с сигналами, близкими к сигналам того или
другого чистого вида. Характерной особенностью
большинства гибридных сигналов была высокая
вариабельность некоторых параметров песни,
например, наблюдался сбой в чередовании разных
элементов, длительность и амплитуда одного и
того же элемента сильно варьировала (Рис. 2).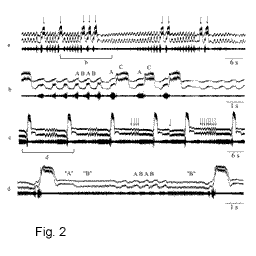 Во время С элемента
некоторые гибридные самцы демонстрировали взмах
задними ногами, похожий на таковой у Ch. oschei,
однако этот демонстрационный сигнал имел
некоторые новые признаки (Рис. 3 c,d).
Во время С элемента
некоторые гибридные самцы демонстрировали взмах
задними ногами, похожий на таковой у Ch. oschei,
однако этот демонстрационный сигнал имел
некоторые новые признаки (Рис. 3 c,d).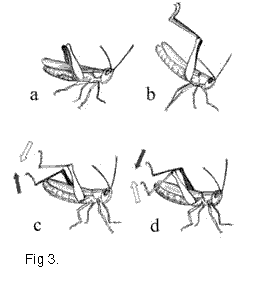 Сравнение гибридных сигналов и сигналов чистых
видов помогло выявить гомологичные элементы в
сигналах Ch. albomarginatus и Сh. oschei (Рис. 4). Так,
элементы В1 и А1 в сигнале Ch. oschei гомологичны
элементам В и А соотвественно.
Сравнение гибридных сигналов и сигналов чистых
видов помогло выявить гомологичные элементы в
сигналах Ch. albomarginatus и Сh. oschei (Рис. 4). Так,
элементы В1 и А1 в сигнале Ch. oschei гомологичны
элементам В и А соотвественно.
В лаборатории были проведены скрещивания двух видов, получены гибриды и записаны сигналы гибридных самцов первого и второго поколений. Анализ сигналов лабораторных гибридов и сравнение с сигналами особей из гибридной зоны показали, что природные гибриды представляют собой преимущественно потомство возвратных скрещиваний (Vedenina et al., 2007).
Встает вопрос, какие
изолирующие механизмы выработаны у Ch. albomarginatus
и Сh. oschei?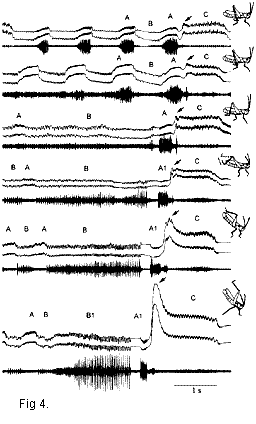 Поведенческие
эксперименты показали, что самки обоих видов в
условиях свободного выбора предпочитают
спариваться с конспецифическими самцами, причем
этот выбор осуществляется главным образом на
основании видоспецифического акустического
сигнала ухаживания (Рис. 6).
Можно заключить, что у данных видов выработаны
этологические изолирующие механизмы, но не
полностью, так как в 10-20% случаев отмечены также
гетероспецифические копуляции.
Поведенческие
эксперименты показали, что самки обоих видов в
условиях свободного выбора предпочитают
спариваться с конспецифическими самцами, причем
этот выбор осуществляется главным образом на
основании видоспецифического акустического
сигнала ухаживания (Рис. 6).
Можно заключить, что у данных видов выработаны
этологические изолирующие механизмы, но не
полностью, так как в 10-20% случаев отмечены также
гетероспецифические копуляции.
Исследование жизнеспособности выращенных в лаборатории гибридов первого и второго поколения показало, что часть гибридов отличалось повышенной эмбриональной и личиночной смертностью (Vedenina et al., 2007). Эти гибриды были потомками самок Ch. oschei. Подобная асимметрия в жизнеспособности реципрокных гибридов описана во многих исследованиях (Harrison, 1983; Orr, 1996; Naisbit et al., 2002). Генетическая несовместимость, которая выражена сильнее в одном направлении скрещиваний, свидетельствует о неполностью установленном изолирующем барьере.
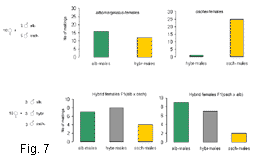
Мы исследовали, насколько гибриды могут конкурировать с родительскими видами за привлечение самок и успешное спаривание. Поведенческие эксперименты показали, что гибридные самки демонстрируют меньшую избирательность, чем самки чистых видов. Самая высокая избирательность отмечена у самок Ch. oschei: в 80-90 % случаев они предпочитали конспецифических самцов (Рис. 7). Самки Ch. albomarginatus не отличали конспецифических от гибридных самцов, но при выборе между конспецифическими самцами и самцами Ch. oschei достоверно предпочитали первых. Гибридные самки не различали три типа самцов и демонстрировали примерно одинаковую частоту копуляций со всеми самцами. Эти результаты свидетельствуют о том, что гибридные самки в смешанных популяциях могут успешно конкурировать с самками чистых видов за привлечение полового партнера. Напротив, гибридные самцы могут конкурировать с самцами Ch. albomarginatus, но проигрывать самцам Ch. oschei.
Мы предполагаем, что существует также биотопическая изоляция между двумя видами в контактной зоне. В настоящее время исследуются биотопические предпочтения Ch. albomarginatus и Сh. oschei.